Sections of the site
Editor's Choice:
- Genetic maps of chromosomes"
- Fumigation of a wooden house against beetles with phosphine
- Visible light spectrum. Visible range. Visible light is an electromagnetic wave
- Hydrogen: distribution in nature
- Interaction of parallel currents figure
- 4 internal friction in liquids
- “About the world of the dead” - T. Tikhoplav. Vitaly and Tatyana Tikhoplav Where did Tatyana Tikhoplav disappear from contact?
- Why does the Moon only attract water?
- Geography What you need to take with you
- How teachers are certified Compliance with the position held for biology teachers
Advertising
|
Genes localized on one chromosome form groups; linkages and are inherited, as a rule, together. The number of linkage groups in diploid organisms is equal to the haploid set of chromosomes. Women have 23 clutch groups, men have 24. The linkage of genes located on the same chromosome can be complete or incomplete. Complete linkage of genes, i.e. joint inheritance, is possible in the absence of the crossing over process. This is typical for genes of sex chromosomes, organisms heterogametic in sex chromosomes (XY, XO), as well as for genes located near the centromere of the chromosome, where crossing over almost never occurs. In most cases, genes localized on one chromosome are not completely linked, and in prophase I of meiosis, identical sections are exchanged between homologous chromosomes. As a result of crossing over, allelic genes that were part of the linkage groups in the parental individuals are separated and form new combinations that enter the gametes. Gene recombination occurs. Gametes and zygotes containing recombinations of linked genes are called crossover. Knowing the number of crossover gametes and the total number of gametes of a given individual, you can calculate the frequency of crossing over as a percentage using the formula: the ratio of the number of crossover gametes (individuals) to the total number of gametes (individuals) multiplied by 100%. The percentage of crossing over between two genes can be used to determine the distance between them. The unit of distance between genes - the morganid - is conventionally taken to be 1% crossing over. The frequency of crossing over also indicates the strength of linkage between genes. The strength of linkage between two genes is equal to the difference between 100% and the percentage of crossing over between these genes. The genetic map of a chromosome is a diagram of the relative arrangement of genes located in the same linkage group. Determining linkage groups and distances between genes is not the minimum stage of constructing a genetic map of a chromosome; cabbage sprat also needs to establish the correspondence of the linkage group being studied to a specific chromosome. The linkage group is determined using the hybridological method, i.e. by studying the results of crossing, and the study of chromosomes - by the cytological method with a microscopic examination of preparations. To determine whether a given linkage group corresponds to a specific chromosome, chromosomes with an altered structure are used. A standard analysis of dihybrid crossing is performed, in which one character under study is encoded by a gene localized on a chromosome with an altered structure, and the second by a gene localized on any other chromosome. If linked inheritance of these two characteristics is observed, we can talk about the connection of this chromosome with a certain linkage group. Analysis of genetic and cytological maps made it possible to formulate the main provisions of the chromosomal theory of heredity. 1. Each gene has a specific constant location (locus) and chromosome. 2. Genes on chromosomes are located in a certain linear sequence. 3. The frequency of crossing over between genes is directly proportional to the distance between them and inversely proportional to the strength of linkage. Block of information. Linkage of genes. At the beginning of the 20th century (1902-1907), the American scientist W. Setton and the German embryologist T. Boveri discovered parallelism in the inheritance of characteristics and the behavior of the chromosomes of the cell nucleus during the process of gametogenesis and fertilization. This confirmed the localization of hereditary information in chromosomes. It has been established that the number of genes significantly exceeds the number of chromosomes. So a person has 46 chromosomes, and there are from 70,000 to 100,000 genes. Consequently, a large number of genes are localized on each chromosome. Genes on the same chromosome are inherited together (linked). An experimental study of this phenomenon was carried out by the American geneticist T. Morgan and his collaborators: A. Sturtevant, A. Bridges and G. Möller in 1910-1916. These studies confirmed the chromosomal localization of genes and formed the basis of the chromosomal theory of heredity. Basic provisions of the chromosomal theory of Heredity.1. Each gene occupies a specific place on the chromosome - a locus. 2. Genes on a chromosome are arranged linearly in a certain sequence. 3. Different chromosomes contain different numbers of genes. The set of genes on each non-homologous chromosome is unique. 4. Genes of one chromosome form a “linkage group and are inherited together, i.e. linked. 5. The number of linkage groups is equal to the number of chromosomes in the haploid set (there are four in Drosophila, 10 in corn, 20 in mice, 23 in humans). 6. An exchange of allelic genes, i.e. crossing over, can occur between homologous chromosomes. 7. The frequency of crossing over is directly proportional to the distance between genes in the linkage group. 8. A special unit, the morganide (M), is taken as the unit of distance between genes in a linkage group. 1M=1% crossing over. There are complete and incomplete linkage of genes. Full grip. In experiments on Drosophila it was found that the development traits that are inherited linked and controlled by genes of one chromosomes. Genes for body color (b - gray and B - black) and wing length (v - normal and V - short, vestigial wings) are localized in one pair of homologous chromosomes. Crossing gray flies with normal wings and gray flies with vestigial wings produces gray hybrids with normal wings in the first generation. When carrying out the analyzing crossing, males were selected from p, since it is known that male Drosophila have achiasmatic spermatogenesis (i.e., crossing over does not occur and the completeness of gene linkage is not disturbed in any way). As a result of such crossing, individuals of two phenotypes similar to the original parental forms, and in equal quantities: 1:1 phenotypic splitting. Considered together, the results of both crossings convince us that the development of the analyzed traits is controlled by different genes, and linked inheritance is explained by the localization of genes on the same chromosome. The completeness of adhesion in this case is not disturbed by anything. This gene linkage is complete. To study incomplete linkage from P, females (genotype B||b) were selected; crossing over occurs in C females during gametogenesis. Therefore, a diheterozygous individual forms additional, i.e. crossover varieties of gametes. The probability of their formation is determined by the probability of crossing over, i.e. depends on the distance between genes in the linkage group. Not recombinant individuals; y Recombinant individuals; at they are observed to have the same formation of their genotypes combinations, like the original ones, involved crossover parental forms of the gamete. In this example, the phenotypic breakdown in the offspring was as follows: gray flies with long wings - 41.5%; black with short wings - 41.5%; gray shortwings - 8.5%; black longwings - 8.5%. Thus, the probability of the appearance of recombinant individuals in the offspring is 17%. Therefore, the distance between genes B and V in the linkage group is 17 morganiids. The predominance of gray long-winged and black short-winged flies in the offspring indicates that genes B and V; b and v are indeed linked. On the other hand, the appearance of recombinant individuals indicates that in a certain number of cases there is a break in the linkage between genes B and V and genes B and V. This is the result of crossing over. An example of complete linkage of genes in humans is the inheritance of the Rh factor. It is caused by three pairs C, D, K, closely linked to each other, therefore inheritance of Rhesus affiliation occurs according to the type of monohybrid crossing. Another example of the close linkage of genes in humans is the inheritance of cataracts and polydactyly. The genes for hemophilia and color blindness are localized on the X chromosome at a distance of 9.8 morganids (M), i.e. undergo crossing over and are therefore inherited as incompletely linked. Autosomal genes for the Rh factor and the shape of erythrocytes, located at a distance of 3 M from each other, are also an example of incomplete linkage. In 1909, F. Janssens, while studying meiosis in amphibians, discovered chiasmata (crossovers) of chromosomes, which are cytological evidence of crossing over. Since that time, many attempts have been made to explain the mechanism of this phenomenon. There are several theories of crossing over. The most common are two hypotheses. Task No. 4. A tall sweet pea plant with green, wrinkled seeds is crossed with a plant that has dwarf growth and green, round seeds. The offspring were split: 3/4 tall plants with green round seeds and 1/4 tall plants with yellow round seeds. Determine the genotypes of the original plants and F 1 hybrids. Given: A- high growth Genotypes P and F 1 – ? Solution Based on the nature of the splitting of traits in hybrids, we draw conclusions about the nature of the genes and the traits they encode. For a pair of genes that determine plant height, the tall plant was homozygous, since among the hybrids there is no segregation for this trait. For a pair of genes that determine seed color, both plants were heterozygous, since among hybrids for this trait there is a 3:1 split. The parent plant with round seeds was a dominant homozygote, since all hybrids had round seeds.
Answer III. Homework Solve a genetic problem. Chickens have feathered legs ( A) dominate naked ( A), rose-shaped comb ( IN) – over a simple ( b), white plumage ( WITH) – above the colored ( With). A hen with feathered legs, a pink comb and white plumage is crossed with a rooster of the same type. Among the offspring was a chicken with bare legs, a simple comb and colored feathers. Determine the genotypes of the parents. Lesson No. 12–13. Linked inheritance of genes. Full and incomplete clutch. Genetic maps Equipment: tables on general biology illustrating the linked inheritance of genes and traits. During the classes I. Test of knowledge Checking the solution to the problem at home. Given: A– feathered legs Genotypes P –? Solution When crossing two parental individuals, each of which was a carrier of three dominant traits, a carrier of three recessive traits appears among the hybrids. This indicates triheterozygosity of each parent. (31 op., pink, white: 9 gol., pink, white: 3 op., inc., white: 5 op., inc., white: 9 op., pink, painted: 3 op., etc., painted: 3 heads, pink, painted: 1 head, straight, dyed.) Answer: Independent work Each student receives a card with the text of a genetic problem and solves it in a notebook. Task No. 1. Tomato plants of the Golden Beauty variety have yellow fruits and tall growth, while the Dwarf variety is short with red fruits. How can you get a homozygous dwarf variety with yellow fruits using these varieties? Task No. 2. Tomato fruits are red and yellow, smooth and fluffy. The red gene is dominant, the furry gene is recessive. Both pairs are on different chromosomes. What kind of offspring can be expected from crossing heterozygous tomatoes with an individual homozygous for both recessive genes? Task No. 3. Tomato fruits are red and yellow, smooth and fluffy. The red gene is dominant, the furry gene is recessive. Both pairs are on different chromosomes. Of the tomatoes harvested on the collective farm, there were 36 tons of smooth red tomatoes and 12 tons of red fluffy ones. How many yellow fluffy tomatoes will be in the collective farm harvest if the starting material was heterozygous for both traits? Task No. 4. In cattle, the polled gene dominates the horned gene, and the black gene dominates the red gene. Both pairs of genes are not linked, that is, they are located in different pairs of chromosomes. On the breeding farm, black polled cows were crossed with a black polled bull for a number of years. 896 heads of young animals were obtained, of which 535 were black polled calves and 161 were red polled calves. How many horned calves were there and how many of them were red? Task No. 5. In cattle, the polled gene dominates the horned gene, and the black gene dominates the red gene. Both pairs of genes are not linked, that is, they are located in different pairs of chromosomes. The farm produced 984 calves from 1,000 horned red cows. Of these, 472 are red, polled - 483, horned - 501. Determine the genotypes of the parents and the percentage of black calves. Task No. 6. In humans, the gene for brown eyes dominates the gene for blue eyes, and the ability to use predominantly the right hand dominates left-handedness. Both pairs of genes are located on different chromosomes. What kind of children can they be if the father is left-handed, but heterozygous for eye color, and the mother is blue-eyed, but heterozygous for the ability to use her hands? Task No. 7. In humans, the gene for brown eyes dominates the gene for blue eyes, and the ability to use predominantly the right hand dominates left-handedness. Both pairs of genes are located on different chromosomes. A blue-eyed right-hander married a brown-eyed right-hander. They had two children: a brown-eyed left-hander and a blue-eyed right-hander. Determine the probability of birth in this family of blue-eyed children who control predominantly the left hand. Task No. 8. Crossing phlox plants with white funnel-shaped flowers with a plant with cream saucer-shaped flowers produced 96 plants with white saucer-shaped flowers. Determine the genotypes of the original plants if it is known that each of the traits is inherited monogenic and the traits are inherited independently. What signs are dominant? Task No. 9. From crossing two white-flowered phlox plants with saucer-shaped flowers in F 1, the following were obtained: 49 plants with white saucer-shaped flowers, 24 with white funnel-shaped flowers, 17 with cream saucer-shaped flowers and 5 with cream funnel-shaped flowers. Based on the results of this cross, is it possible to determine how these traits are inherited? Determine the genotypes of the original plants. What splitting would occur if the original plants were crossed with a plant with cream funnel-shaped flowers from F 1? Task No. 10. When two tomato plants with red bilocular fruits were self-pollinated, one of them produced only plants with red bilocular fruits, and the second produced 24 plants with red bilocular fruits and 10 plants with red multilocular fruits. Is it possible to determine the genotypes of the original plants? What crosses are needed to test your hypothesis? II. Learning new material Linked inheritance of genes G. Mendel traced the inheritance of seven pairs of traits in peas. Many researchers, repeating Mendel's experiments, confirmed the laws he discovered. It was recognized that these laws were of a universal nature. However, in 1906, English geneticists W. Batson and R. Pennett, crossing sweet pea plants and analyzing the inheritance of pollen shape and flower color, discovered that these characters do not give independent distribution in the offspring. Descendants always repeated the characteristics of their parent forms. Gradually, evidence of exceptions to Mendel's third law accumulated. It became clear that not all genes are characterized by independent distribution in the offspring and free combination. Any organism has a variety of morphological, physiological, biochemical and other characteristics and properties, and each characteristic or property is controlled by one or more genes localized in the chromosomes. However, if the number of genes in an organism is huge and can amount to tens of thousands, then the number of chromosomes is relatively small and, as a rule, is measured in several tens. Therefore, hundreds and thousands of allelic genes forming linkage groups are localized in each pair of chromosomes. A complete correspondence has been established between the number of linkage groups and the number of chromosome pairs. For example, corn has a set of chromosomes 2n = 20 and 10 linkage groups, and Drosophila has 2n = 8 and 4 linkage groups, that is, the number of linkage groups is equal to the haploid set of chromosomes. Thomas Morgan Law Genes localized on the same chromosome are transmitted together, and their mode of inheritance differs from the inheritance of genes localized in different pairs of homologous chromosomes. So, for example, with an independent distribution of chromosomes, a dihybrid AaBb forms four types of gametes ( AB, aB, Ab, ab), and under the condition of complete linkage, the same dihybrid will produce only two types of gametes ( AB And ab), since these genes are located on the same chromosome. The development of the problem of linked inheritance of genes belongs to the school of T. Morgan (1866–1945). If Mendel conducted his experiments on peas, then for Morgan the main object was the fruit fly Drosophila. The fly produces numerous offspring every two weeks at a temperature of 25 °C. The male and female are clearly distinguishable - the male has a smaller and darker abdomen. In addition, they differ in numerous characteristics and can reproduce in test tubes on a cheap nutrient medium. Studying patterns of inheritance genes localized on the same chromosome, Morgan concluded that they are inherited linked. This is T. Morgan's law. Full and partial clutch To determine the type of inheritance of two pairs of genes (linked or independent), it is necessary to conduct an analytical crossing and, based on its results, draw a conclusion about the nature of gene inheritance. Let us consider three possible variants of the results of an analyzing cross. 1) Independent inheritance.
If, as a result of analytical crossing, four classes of phenotypes are formed among hybrids, it means that the genes are inherited independently. 2) Complete gene linkage.
With complete gene linkage A And IN according to the results of the analyzing crossing, they find - 3) Incomplete gene linkage.
In case of incomplete linkage of genes A And IN When analyzing crosses, four phenotypes appear, two of which have a new combination of genes: Ab‖ab; aB‖ab. The appearance of such forms indicates that the dihybrid with gametes AB│ and ab│ forms crossover gametes Ab│ and aB│. The appearance of such gametes is possible only as a result of the exchange of sections of homologous chromosomes, that is, in the process of crossing over. The number of crossover gametes is significantly smaller than non-crossover gametes. The crossover frequency is proportional to the distance between genes. The closer the genes are located on a chromosome, the closer the linkage between them and the less often they separate during crossover. And vice versa, the farther the genes are from each other, the weaker the linkage between them and the more often they cross over. Consequently, the distance between genes on chromosomes can be judged by the frequency of crossover. Genetic maps Genetic mapping usually means determining the position of a gene in relation to other genes. Let's consider the procedure for compiling genetic maps. 1. Establishment of the linkage group (that is, determination of the chromosome in which the gene is localized). To do this, it is necessary to have at least one marker gene in each linkage group. 2. Finding the location of the gene under study in the chromosome. To do this, the mutant form is crossed with a normal one and the result of crossing over is taken into account. 3. Determination of the distance between linked genes, which makes it possible to draw up genetic maps of chromosomes, which indicate the order of genes in the chromosomes and their relative distances from each other. The higher the crossing over frequency, the greater the distance between the genes. If it is established that between linked genes A And IN crossing over frequency is 10%, and between genes IN And WITH– 20%, then it is obvious that the distance Sun 2 times more than AB. The distance between genes is expressed in units corresponding to 1% crossing over. These units are called morganids. Thus, based on data on the frequency of crossing over, genetic maps are compiled. III. Consolidation of knowledge Solving a genetic problem A female Drosophila heterozygous for the recessive genes for dark body color and miniature wings was crossed with a male who had a dark body and miniature wings. From this crossing the following was obtained: – 244 flies with a dark body and miniature wings; Based on the given data, determine whether these two pairs of genes are linked or not. How are genes linked? Given: A– gray body The nature of gene inheritance A And IN -? Solution The results of splitting among hybrids (two phenotypic classes are dominant and repeat phenotypically and genotypically the parental forms, and the other two classes of phenotypes are represented by a small number of individuals) indicate incomplete linkage of genes A And IN.
Answer: genes A And IN are inherited concatenated; the clutch is incomplete. IV. Homework Study the textbook paragraph (linked inheritance of genes, T. Morgan's law, complete and incomplete linkage, genetic maps). Solve a genetic problem. Determine the frequency of crossing over between genes if, when crossing gray long-winged flies with black short-winged flies in F 1, all flies were gray long-winged, and in the analyzing crossing of F 1 females with a black short-winged male the following was obtained: – 722 gray long-winged flies; Lesson No. 14–15. Chromosomal theory of heredity. Non-chromosomal (cytoplasmic) inheritanceEquipment: tables on general biology illustrating the linked inheritance of genes and traits; portraits of scientists who made a special contribution to the creation of the chromosomal theory of heredity. During the classes I. Test of knowledge 1. Oral test of knowledge on the following questions: Linked inheritance of genes. T. Morgan's Law. 2. Checking the solution to the problem at home. Given: A– gray body Crossover frequency – ? Solution We draw up schemes for the first and second crossings.
N (hybrids) = 722 + 778 + 161 + 139 = 1800 individuals. N (recombinants) = 161 + 139 = 300 individuals. (300 × 100) : 1800 = 16.6% Answer: 16,6%. II. Solving problems in the classroom Task No. 1. If we assume that genes A And IN are linked and the crossover between them is 10%, then what gametes and in what quantity will be formed by the diheterozygote AB‖ab? Solution Crossover between genes A And IN is 10%, this means that a total of 10% of crossover gametes are formed (5% of each type). The share of non-crossover gametes remains 90% (45% of each type).
Answer: AB (45%); ab (45%); Ab (5%); aB (5%). Task No. 2. The smooth shape of corn seeds is dominant over the wrinkled shape, and the colored seeds are dominant over the uncolored ones. Both signs are linked. When crossing corn with smooth, colored seeds with a plant with wrinkled, uncolored seeds, the following offspring were obtained: painted smooth – 4152 individuals; Determine the distance between genes. Given: A– smooth seeds Distance AB – ? Solution Let's draw up a crossing scheme.
2) We count the total number of hybrids: N (hybrids) = 4152 + 4163 + 152 + 149 = 8616 individuals. 3) We count the number of recombinant individuals: N (recombinants) = 152 + 149 = 301 individuals. 4) Determine the crossing over frequency: (301 × 100) : 8616 = 3.5% 5) One percent crossing over is equal to 1 morganid, so the distance between genes A And IN equals 3.5 morganids. Answer: 3.5 morganids. III. Learning new material Chromosomal theory of heredity Thanks to the research of A. Weisman, T. Boveri, T. Morgan and other outstanding geneticists and cytologists, by the 40s. XX century The chromosomal theory of heredity was formulated. The modern chromosomal theory of heredity includes the following postulates: 1) the characteristics of organisms are formed under the influence of genes located on chromosomes; Thus, the chromosomal theory of heredity is an outstanding achievement of biological science. It was the result of combining knowledge gained in two biological disciplines: genetics and cytology. Cytoplasmic inheritance Along with the facts confirming the chromosomal theory of heredity, in the process of the formation of genetics as a science, facts about heredity began to accumulate that did not obey established patterns: inheritance only through the maternal line, deviations from Mendelian numerical relationships, etc.
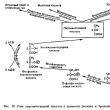
These cases could only be explained by the localization of the genes that determine this trait in the cytoplasm, that is, the cytoplasm also plays a certain role in the formation of certain traits. This phenomenon is called cytoplasmic, or extrachromosomal, heredity. For example, chloroplasts of higher plants during sexual reproduction are transmitted through the maternal line; male gametes do not contain them, therefore, the zygote contains chloroplasts that were in the egg. Chloroplasts contain their own circular DNA, which ensures the synthesis of some proteins and RNA responsible for a number of characteristics. In the night beauty and snapdragon, variegation phenomena associated with mutations in the DNA of some chloroplasts are known. These mutations can cause chloroplasts to lose their green color. When a cell divides, the distribution of chloroplasts among daughter cells occurs randomly; daughter cells may contain colored, colorless, or both chloroplasts. If the egg contained colorless chloroplasts, then an uncolored plant will develop from the zygote; after using up the supply of nutrients, it dies. If the egg contains only green chloroplasts, a normal green plant will develop. If both green and colorless chloroplasts enter the egg, the plant will be variegated. In the same way, traits associated with mutations that occur in mitochondria are inherited. IV. Homework Study the textbook paragraph and notes made in class (provisions of the chromosomal theory of heredity, cytoplasmic heredity). In 1906, W. Batson and R. Punnett, crossing sweet pea plants and analyzing the inheritance of pollen shape and flower color, discovered that these characteristics do not give independent distribution in the offspring; hybrids always repeated the characteristics of the parent forms. It became clear that not all traits are characterized by independent distribution in the offspring and free combination. Each organism has a huge number of characteristics, but the number of chromosomes is small. Consequently, each chromosome carries not one gene, but a whole group of genes responsible for the development of different traits. He studied the inheritance of traits whose genes are localized on one chromosome. T. Morgan. If Mendel conducted his experiments on peas, then for Morgan the main object was the fruit fly Drosophila. Drosophila produces numerous offspring every two weeks at a temperature of 25 °C. The male and female are clearly distinguishable in appearance - the male has a smaller and darker abdomen. They have only 8 chromosomes in a diploid set and reproduce quite easily in test tubes on an inexpensive nutrient medium. By crossing a Drosophila fly with a gray body and normal wings with a fly having a dark body color and rudimentary wings, in the first generation Morgan obtained hybrids with a gray body and normal wings (the gene that determines the gray color of the abdomen dominates the dark color, and the gene that determines development of normal wings, - above the gene of underdeveloped wings). When carrying out an analytical crossing of an F 1 female with a male who had recessive traits, it was theoretically expected to obtain offspring with combinations of these traits in a ratio of 1:1:1:1. However, in the offspring, individuals with characteristics of the parental forms clearly predominated (41.5% - gray long-winged and 41.5% - black with rudimentary wings), and only a small part of the flies had a combination of characters different from those of the parents (8.5% - black long-winged and 8.5% - gray with rudimentary wings). Such results could only be obtained if the genes responsible for body color and wing shape are located on the same chromosome.
1 - non-crossover gametes; 2 - crossover gametes. If the genes for body color and wing shape are localized on one chromosome, then this crossing should have resulted in two groups of individuals repeating the characteristics of the parental forms, since the maternal organism should form gametes of only two types - AB and ab, and the paternal one - one type - ab . Consequently, two groups of individuals with the genotype AABB and aabb should be formed in the offspring. However, individuals appear in the offspring (albeit in small numbers) with recombined characters, that is, having genotypes Aabb and aaBb. In order to explain this, it is necessary to recall the mechanism of formation of germ cells - meiosis. In the prophase of the first meiotic division, homologous chromosomes are conjugated, and at this moment an exchange of regions can occur between them. As a result of crossing over, in some cells, sections of chromosomes are exchanged between genes A and B, gametes Ab and aB appear, and, as a result, four groups of phenotypes are formed in the offspring, as with the free combination of genes. But, since crossing over occurs during the formation of a small part of gametes, the numerical ratio of phenotypes does not correspond to the ratio 1:1:1:1. Clutch group- genes localized on the same chromosome and inherited together. The number of linkage groups corresponds to the haploid set of chromosomes. Chained inheritance- inheritance of traits whose genes are localized on the same chromosome. The strength of linkage between genes depends on the distance between them: the further the genes are located from each other, the higher the frequency of crossing over and vice versa. Full grip- a type of linked inheritance in which the genes of the analyzed traits are located so close to each other that crossing over between them becomes impossible. Incomplete clutch- a type of linked inheritance in which the genes of the analyzed traits are located at a certain distance from each other, which makes crossing over between them possible. Independent inheritance- inheritance of traits whose genes are localized in different pairs of homologous chromosomes. Non-crossover gametes- gametes during the formation of which crossing over did not occur. Non-recombinants- hybrid individuals that have the same combination of characteristics as their parents. Recombinants- hybrid individuals that have a different combination of characteristics than their parents. The distance between genes is measured in Morganids— conventional units corresponding to the percentage of crossover gametes or the percentage of recombinants. For example, the distance between the genes for gray body color and long wings (also black body color and rudimentary wings) in Drosophila is 17%, or 17 morganids. In diheterozygotes, dominant genes can be located either on one chromosome ( cis phase), or in different ( trans phase).
1 - Cis-phase mechanism (non-crossover gametes); 2 - trans-phase mechanism (non-crossover gametes). The result of T. Morgan's research was the creation of chromosomal theory of heredity:
Go to lectures No. 17“Basic concepts of genetics. Mendel's laws" 32. Peculiarities of inheritance of a trait with incomplete and complete couplingLINKED INHERITANCE OF CHARACTERISTICS We became acquainted with dihybrid crossing and realized that the independent combination of traits is explained by the fact that the splitting of one pair of allelic genes that determine the corresponding traits occurs independently of the other pair. However, this is observed only in the case when the genes of different pairs are located in different pairs of chromosomes and when the hybrid germ cells are formed in meiosis, the paternal and maternal chromosomes are independently combined. But the number of chromosomes is very limited compared to the number of characters, each of which develops under the control of a specific gene. Thus, about 7000 genes are known in Drosophila with four pairs of chromosomes. It is assumed that a person has at least 50 thousand genes with 23 pairs of chromosomes, etc. It follows that hundreds of alleles must be localized in each pair of chromosomes. Naturally; that there is linkage between genes that are located on the same chromosome and when germ cells are formed, they must be transmitted together. Linked inheritance was discovered in 1906 by English geneticists W. Batson and R. Punnett while studying the inheritance of traits in sweet peas, but they were unable to reveal the reasons for this phenomenon. The nature of linked inheritance was clarified in 1910 by scientists T. Morgan and his collaborators K. Bridges and A. Sturtevant. As an object of study, they chose the fruit fly Drosophila, which turned out to be very convenient for genetic experiments. The cells of the Drosophila body contain 4 pairs of chromosomes. It is distinguished by very high fertility - one pair produces more than a hundred offspring. It has a high development speed - within 12-15 days after fertilization, a larva, pupa and an adult develop from the egg, which is almost immediately capable of producing offspring. More than twenty generations can be studied over the course of a year. Flies are gray in color, with red eyes. are small in size (about 3 mm), easily diluted in biological test tubes; You can use magnifying glasses to study their signs. When looking at hundreds of thousands of individuals, Morgan discovered many different mutations: there were flies with black and yellow bodies, with white and other colored eyes, with altered shape and position of wings, etc. Sometimes there were individuals with several mutations at once, for example black body, rudimentary wings, cinnabar eyes. Studying the inheritance of different pairs of characters during dihybrid and polyhybrid crossings, Morgan and his colleagues discovered a large number of examples of their linked (joint) inheritance. All studied characters were divided into four linkage groups in accordance with the number and size of chromosomes in Drosophila. On this basis, Morgan concluded that the genes that determine these traits are located on chromosomes. Genes located on the same chromosome constitute a linkage group. Gene linkage is the joint inheritance of genes located on the same chromosome. The number of linkage groups corresponds to the haploid number of chromosomes. For example, Drosophila has 4 linkage groups, humans have 23, cattle have 30, pigs have 19, etc. The idea of the location of genes in chromosomes was expressed by Setton back in 1902. He discovered parallelism in the behavior of chromosomes in meiosis and the inheritance of traits in one of the grasshopper species. Further research conducted by Morgan showed that the linkage of genes located on the same chromosome can be complete or incomplete. The difference in the behavior of linked and independently inherited genes is most clearly revealed when conducting an analytical cross. With independent inheritance of two pairs of characters in a hybrid Fi (АаББ) 4 types of gametes are formed with equal probability: AB,Ab, aB, ab. When crossed with a complete recessive (aabb) the number of varieties of gametes in a hybrid determines the number of types of descendants and the same probability of their appearance, since the gametes of a recessive individual (ab) cannot change the manifestations of dominant and recessive genes of the hybrid gametes. As a result, the ratio of offspring phenotypes will be 1:1:1:1. If both pairs of allelic genes are located in one pair of chromosomes, then during the formation of germ cells the genes of these alleles will not be able to combine freely. In this case, linked inheritance is observed. FULL CLUTCH T. Morgan crossed black long-winged females with gray males with rudimentary wings. In Drosophila, gray body color dominates over black, and long wings dominate over rudimentary wings. Let us denote the gray body color gene IN, allelic gene for black body color b; Longwing™ gene V, allelic to it is the gene for rudimentary wings v. Both pairs of these genes are located on the same second pair of chromosomes. For both pairs of Characters, the parental forms were homozygous: female for the recessive trait of black body (bb) and the dominant trait long-winged™ (V.V.), male by dominant gray coloration. (BB) and the recessive trait of rudimentary wings (w>. The gametes of the parents, during reduction division, receive from the maternal form a chromosome with genes b And V, from the father's - with genes IN and v. All offspring of the first generation (Fi) had a gray body and long wings (Fig. 13) and were heterozygous for both pairs of traits (bV/ Bv). Then males were selected from Fi and crossed with females homozygous for both recessive genes, black rudiments (bv/ bv), i.e., an analytical crossing was carried out, as a result of which, with an independent combination of characters, the offspring of four phenotypes should have been obtained in equal proportions: gray long-winged, gray with rudimentary wings, black long-winged, black with rudimentary wings, but the descendants of only two phenotypes were obtained , similar to the original parental forms: black long-winged and gray short-winged. In this case, complete concatenation of characteristics is observed. This is due to the fact that in a heterozygous male, both the gene for black coloration and the gene for long wings are located on the same chromosome from a homologous pair, and the gene for gray coloration and the gene for rudimentary wings are located on the other. During spermiogenesis during meiosis, homologous chromosomes separate into different germ cells. Only two varieties of gametes are produced: one with a chromosome that carries genes Kommersant And V, the other with the chromosome on which the genes are located IN and v. When these gametes are combined with gametes of an individual with recessive traits, only two types of offspring are formed. In complete linkage, genes located on the same chromosome are always transmitted together. Complete linkage has so far been established only in fruit flies and female silkworms. INCOMPLETE CLUTCH In the next experiment, as in the previous one, Morgan crossed black long-winged females with gray rudimentary-winged males. In the first generation, all the offspring were gray long-winged. Then he again made an analytical cross, but from the first generation he selected not a male, but a female and crossed her with a black male with rudimentary wings (Fig. 14). In this case, the offspring appeared not of two types, as with complete linkage, but of four: gray with rudimentary wings, black long-winged, gray long-winged and black with rudimentary wings, but not in equal proportions, as with an independent combination of characters, but with a significant predominance phenotypes similar to the parental forms. 41.5% of the flies were gray with rudimentary wings, like one original parent, and 41.5% were black long-winged, like the other original parent. Only 17% of the offspring were born with a new combination of characteristics: 8.5% were black with rudimentary wings and 8.5% were gray long-winged. Thus, 83% of the descendants had a combination of characteristics like the original parental forms, but individuals also appeared with a new combination of characteristics. Therefore, the clutch is incomplete./ The question arose: why did individuals with a new combination of parental characteristics appear? To explain this phenomenon, Morgan used and developed the theory chiasmotypes Belgian cytologist Janssens. In 1909, Janssens observed that during spermiogenesis in the salamander in the prophase of meiosis, homologous chromosomes conjugate, and then, at the beginning of divergence, form figures in chromosomes exchange sections. If linked genes lie on the same chromosome and recombination of these genes occurs in heterozygotes during the formation of gametes, it means that homologous chromosomes exchanged their parts during meiosis. The exchange of homologous chromosomes with their parts is called decussation or cross-singover(the English word crossingover means the formation of a cross). Individuals with new combinations of characters resulting from crossing over are called crossovers. Let's return to Figure 14. In female Fi, heterozygous for both pairs of traits, genes are located on one of the homologous chromosomes Kommersant And V, in the other - genes allelic to them IN and v. In the prophase of reduction division, when two homologous chromosomes are united into one bivalent, each chromosome is doubled and consists of two chromatids. There will be 4 chromatids in total. Between two chromatids of homologous chromosomes, an exchange of their parts occurs. As a result, the gene Kommersant, located in the chromatid of one homologous chromosome, can connect with the v gene located in the chromatid of another homologous chromosome, and as a result of one event, a second chromatid is formed, where the genes are connected IN And V. Subsequently, the chromatids will separate and crossover gametes with chromosomes with a new combination of genes will be formed. (bv And B.V.). The other two chromatids from a pair of homologous chromosomes do not participate in the crossover and retain the maternal ones in the original combination (bV) and paternal (Bv) genes. The formation of new crossover gametes ensured the appearance of fruit flies with a new combination of characters: black with rudimentary wings and gray long-winged ones. However, most of the offspring will be similar to the original parents (black long-winged and gray short-winged). Morgan concludes that the number of appearances of new forms depends on the frequency of crossover, which is determined by the following formula: If, for example, the total number of descendants is 900, and the new crossover forms are 180, then the crossover frequency will be 20 %. Morgan found that frequency of crossover between a specific pair of genes- a relatively constant value, but different for different pairs of genes. Based on this, it was concluded that The frequency of crossover can be used to judge the distances between genes. The unit of measurement of the cross is taken to be equal to 1%. Sometimes she is called Morganida. The amount of crossover depends on the distance between the genes being studied. The more distant the genes are from each other, the more often crossover occurs; the closer they are located, the less likely it is to cross. It has been established that the number of crossover individuals to the total number of descendants never exceeds 50%, since at very large distances between genes, double crossing over occurs more often and some crossover individuals remain unaccounted for. They can be taken into account when studying not two pairs of linked traits, but three or four. In this case, taking into account double and triple crossovers, it is possible to more accurately judge the distances and frequency of crossover between genes. |











| Read: |
|---|
New
- Fumigation of a wooden house against beetles with phosphine
- Visible light spectrum. Visible range. Visible light is an electromagnetic wave
- Hydrogen: distribution in nature
- Interaction of parallel currents figure
- 4 internal friction in liquids
- “About the world of the dead” - T. Tikhoplav. Vitaly and Tatyana Tikhoplav Where did Tatyana Tikhoplav disappear from contact?
- Why does the Moon only attract water?
- Geography What you need to take with you
- How teachers are certified Compliance with the position held for biology teachers
- Certificate of basic general education What is the series in the certificate




